Glycine and the NMDA Receptor: The Co-Agonist Behind Learning and Memory
Roon Team
Glycine and the NMDA Receptor: The Co-Agonist Behind Learning and Memory
Most people file glycine under "calming amino acid." It is, in the spinal cord and brainstem, where it acts as a major inhibitory neurotransmitter. But the same molecule does something almost opposite in your forebrain, and that second job is the one tied to how you learn and remember.
The glycine NMDA receptor relationship is one of the stranger setups in neuroscience. Glycine doesn't fire the NMDA receptor on its own. It hands the receptor a permission slip, and only when a second signal arrives does anything happen.
That two-key design is not an accident. It is the molecular reason your brain can tell the difference between background noise and a memory worth keeping.
Key Takeaways
- The NMDA receptor needs two chemical signals to open: glutamate and a co-agonist (glycine or D-serine). One alone does nothing.
- Glycine binds the GluN1 subunit at what scientists call the "glycine site," while glutamate binds GluN2.
- This dual requirement makes the NMDA receptor a coincidence detector, the cellular basis of long-term potentiation (LTP) and learning.
- Glycine is both an inhibitory neurotransmitter (spinal cord) and an excitatory co-agonist (forebrain). Same molecule, two roles.
- The glycine site is an active drug target for cognition, especially in schizophrenia research.
What the NMDA Receptor Actually Does
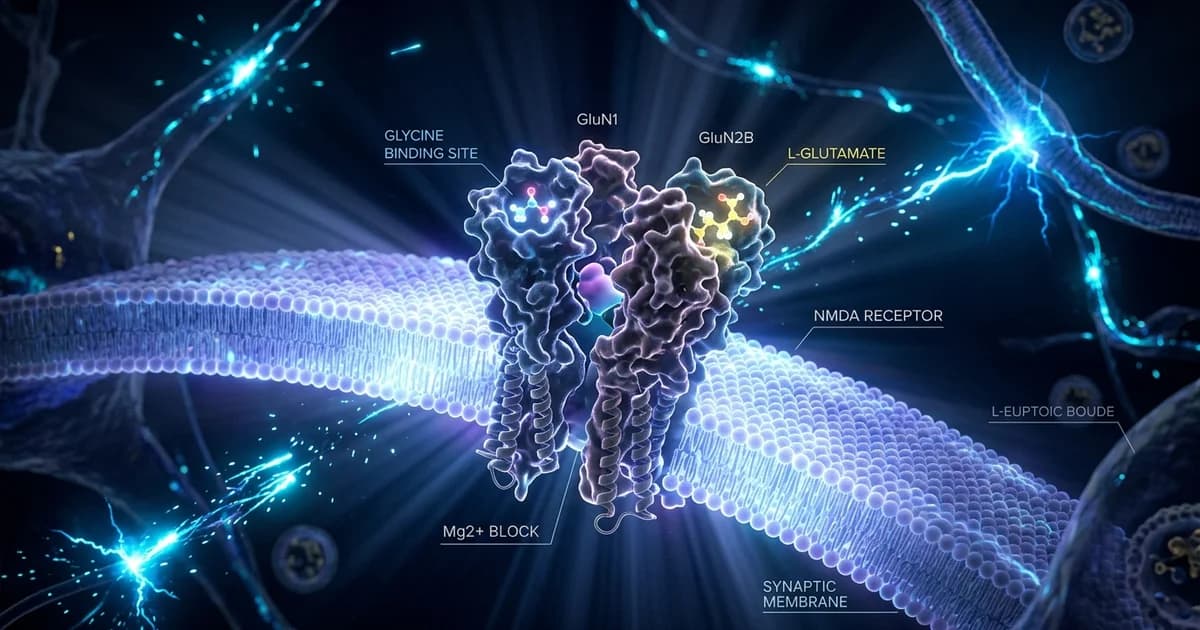
The NMDA receptor is a gate for calcium and sodium into a neuron, and it is famously hard to open. N-methyl-d-aspartate (NMDA) receptors are Hebbian-like coincidence detectors, requiring binding of glycine and glutamate in combination with the relief of voltage-dependent magnesium block to open an ion conductive pore across the membrane bilayer.
Read that again, because three things have to line up at once.
First, glutamate has to bind. Second, a co-agonist (glycine or D-serine) has to be bound. Third, the cell has to already be depolarized enough to kick out a magnesium ion that physically plugs the channel. The binding of the ligands is typically not sufficient to open the channel as it may be blocked by Mg2+ ions which are only removed when the neuron is sufficiently depolarized.
Only when all three conditions overlap does calcium pour in. That calcium signal is what tells the neuron a connection just mattered.
This is why the receptor is built the way it is. It refuses to respond to any single input. It waits for a coincidence.
Why Glycine Is the Co-Agonist, Not the Trigger
Glutamate is the "what happened" signal. Glycine is the "are we ready to listen" signal.
The NMDA receptor is a four-part protein, and the parts don't all listen to the same chemical. N-methyl-D-aspartate receptors (NMDARs) are ionotropic glutamate receptors built as heterotetramers composed of GluN1 (with eight splice variants), GluN2 (GluN2A through GluN2D), and/or GluN3 (GluN3A and GluN3B) subunits.
Glutamate docks onto the GluN2 subunits. Glycine docks onto GluN1. Both binding events are required at the same time, which is what the term glycine co-agonist captures. At central nervous system synapses, agonist binding to postsynaptic ionotropic glutamate receptors results in signaling between neurons. NMDA receptors are a unique family that activate in response to the concurrent binding of glutamate and glycine.
So glycine never pulls the trigger. It loads the gun. Without glycine sitting in its pocket, glutamate can scream all it wants and the channel stays shut.
Glycine, D-Serine, and the Glycine Site
Here is a twist that surprises most people: glycine may not even be the main co-agonist at many synapses. D-serine often is.
The two molecules compete for the same spot, the glycine site on GluN1. In some brain regions, the brain seems to prefer one over the other depending on the type of signal. In hippocampus and amygdala, D-serine is thought to serve as the endogenous agonist responsible for tonic activation mediating NMDAR-mediated currents in response to low levels of synaptic activity.
Research suggests D-serine took over a role once assumed to belong to glycine. By binding to the NMDAR co-agonist site, endogenous D-serine could support NMDAR-dependent synaptic plasticity, a common type of use-dependent changes in synaptic connections thought to represent a cellular mechanism of learning. D-serine could therefore play the role previously attributed to the NMDAR co-agonist glycine.
The two are linked at the source. Astrocytes, the support cells around neurons, run a serine shuttle that feeds the system, and D-serine regulates several NMDA-dependent processes, plasticity chief among them.
The practical point: when you read "glycine site," think of it as the co-agonist socket. Glycine and D-serine are two keys cut for the same lock.
Glycine, Learning, and Memory
The reason any of this matters for cognition comes down to one process: long-term potentiation.
LTP is the strengthening of a synapse after repeated use, and it depends directly on NMDA receptors. It has been suggested that the activation of the NMDA receptor is required for long-term potentiation (LTP) in the hippocampus, amygdala, and medial septum. This mechanism has been implicated in memory formation.
LTP is widely treated as the cellular shorthand for how memories physically form. NMDA receptor-dependent long-term potentiation is extensively studied since it is believed to use the same molecular mechanisms that are required for many forms of learning and memory.
Now connect the dots. The NMDA receptor drives LTP. The NMDA receptor cannot open without a co-agonist in the glycine site. So glycine learning memory links are not metaphorical. The co-agonist is a hard gating requirement for the receptor that builds memory.
Different subunits tune the process. NMDA receptors containing NR2A subunits are key contributors to hippocampal long-term potentiation induction in adult animals and have therefore been widely implicated in hippocampus-dependent spatial learning.
This is the heart of nmda receptor cognition. Glutamate carries the message, glycine grants access, depolarization confirms timing, and calcium writes the change.
One Molecule, Two Jobs: The Excitatory and Inhibitory Glycine
Glycine has a split personality, and the glycine excitatory inhibitory contrast is real, not a contradiction.
In the lower nervous system, glycine calms things down. Glycine is the major inhibitory neurotransmitter of the anterior horn of the spinal cord and of brainstem motor nuclei. It opens chloride channels through its own dedicated glycine receptor, which is a separate protein from the NMDA receptor.
In the forebrain, the same amino acid flips function. In addition to its inhibitory postsynaptic action, glycine also acts as an excitatory transmitter. First, it serves as a co-agonist of the NMDA-subtype of excitatory receptors.
So the location and the receptor decide the outcome. Bind the glycine receptor, you inhibit. Bind the glycine site on the NMDA receptor, you help excite.
| Property | Glycine as inhibitory transmitter | Glycine as NMDA co-agonist |
|---|---|---|
| Main location | Spinal cord, brainstem | Forebrain, hippocampus, cortex |
| Receptor | Glycine receptor (GlyR) | NMDA receptor (GluN1 subunit) |
| Ion effect | Opens chloride channels (calms) | Helps open Ca2+/Na+ channel (excites) |
| Functional role | Motor control, reflex tone | Synaptic plasticity, learning |
| Acts alone? | Yes | No, needs glutamate present |
The Glycine Site as a Drug Target
Because the glycine site controls NMDA receptor activity, it has long attracted researchers chasing cognitive treatments.
The strongest interest comes from schizophrenia work, where NMDA receptor underactivity is one leading model. There is convincing evidence that the glycine modulatory site on the NMDA receptor is a valid therapeutic target for improving cognition and associated negative symptoms in schizophrenia.
Clinical attempts to feed the system directly have shown mixed but real signals. According to a 2024 review in the International Journal of Molecular Sciences, the studies varied in sample size and glycine dosage, but the majority reported marked improvements in negative symptoms and cognitive deficits of schizophrenia with glycine treatment.
This is mechanism research, not a recommendation. Glycine is being studied as a tool for understanding the receptor, and dosing in those trials is far beyond anything in food.
Frequently Asked Questions
Is glycine excitatory or inhibitory?
Both, depending on where it acts. In the spinal cord and brainstem, glycine binds its own glycine receptor and works as a major inhibitory neurotransmitter, calming motor circuits. In the forebrain, the same molecule binds the glycine site on the NMDA receptor and acts as an excitatory co-agonist. The receptor and the brain region decide the outcome, not the molecule itself.
What does it mean that glycine is a co-agonist?
A co-agonist is a second required signal. The NMDA receptor will not open from glutamate alone. It needs glycine (or D-serine) bound to a separate site at the same time. Glycine does not trigger the receptor, it grants permission for glutamate to do so. Both binding events must overlap, which is why the receptor is described as needing two keys.
Where does glycine bind on the NMDA receptor?
Glycine binds the GluN1 subunit, often called the glycine site or co-agonist site. Glutamate binds a different subunit, GluN2. Since an NMDA receptor is built from multiple subunits, it can sense both signals at once. This split design is what allows the receptor to require simultaneous inputs before opening its calcium channel.
What is the difference between glycine and D-serine here?
They compete for the same glycine site on GluN1. Glycine was identified first, but later research found that D-serine is the main co-agonist at many synapses, especially in the hippocampus and amygdala. Astrocytes help supply both through a serine shuttle. Functionally, the receptor accepts either as the co-agonist key.
How does the NMDA receptor relate to memory?
The NMDA receptor drives long-term potentiation, the strengthening of synapses that researchers treat as a cellular basis of learning and memory. Because the receptor cannot open without a co-agonist in the glycine site, glycine and D-serine are part of the gating that allows memory formation at the synaptic level. No co-agonist, no NMDA current, weaker plasticity.
Why is the NMDA receptor called a coincidence detector?
Because it only opens when several conditions happen at the same time: glutamate bound, co-agonist bound, and the neuron already depolarized enough to remove a magnesium block. This makes the receptor a built-in timing check. It fires only when an input arrives while the cell is already active, linking events that occur together. That overlap is the physical logic behind associative learning.
Does eating glycine boost memory?
That is not established. The NMDA receptor mechanism explains why the glycine site matters inside the brain, but dietary glycine and clinical co-agonist dosing are different questions with different evidence. The clinical studies on cognition used controlled doses in specific patient groups. Treat the receptor biology as an explanation of how the system works, not as a supplement claim.
The Two-Key Logic Behind a Memory
The NMDA receptor is one of the clearest examples of biology refusing to act on a single input. It demands glutamate, a co-agonist in the glycine site, and the right electrical moment, all at once. Miss any one, and the channel stays shut.
That stubbornness is the point. It lets a neuron ignore noise and respond only when signals coincide, which is exactly what learning requires. Glycine, the molecule we usually call calming, turns out to be a quiet gatekeeper for the receptor that writes memory.
Glutamate gets the headlines. The co-agonist makes the call on whether anyone listens.
Where This Fits in the Bigger Picture of Focus and Cognition
This article stays at the level of pure mechanism, and that is on purpose. Understanding how the NMDA receptor gates learning is foundational neuroscience, separate from any product or protocol.
If you found this useful, it sits in our broader neuroscience explainers series, where we break down the systems behind attention, memory, and focus in plain language. We cover the receptors, the neurotransmitters, and the everyday inputs that shape cognition, without the hype.
Roon itself is a zero-nicotine sublingual focus pouch, not a glycine supplement and not a treatment for any condition. These explainers sit alongside it for one reason: to explain the brain accurately enough that you can reason about it yourself.
Written by Roon Team